
This multiyear trial was conducted at the Scotts Miracle-Gro Research and Development Center, Marysville, Ohio, from 2019 to 2021. Photo by Cassie Reed
One of the major stresses that impacts turfgrass cultivation worldwide is that caused by shade. Shade stress can originate from various sources and can be classified as temporary, like that from trees or vegetation, or permanent, caused by buildings or structures. It is estimated that worldwide 25% of all turfgrass is grown under some form of shade (2, 3). Turfgrass that is cultivated under shade conditions typically has thinner stands, weaker plants that are more susceptible to other stresses and overall lower quality turf.
Among the different cool-season turfgrass species, there are differences in shade tolerance. Most of the species in the fescue family (Festuca species) generally are considered to be the most shade tolerant, while Kentucky bluegrass (KBG) (Poa pratensis) generally is considered the least shade tolerant. Limited work has been done to quantify the minimum daily light integral requirements (DLIm) for bluegrass. In 2015, Meeks et al. conducted a short-term greenhouse study looking at the DLI requirements for Rebel Exceda tall fescue (Festuca arundinacea), two KBG cultivars and five bluegrass hybrids (P. arachnifera × P. pratensis). The KBG cultivar Kenblue was found to have a DLIm of 24.1 moles per square meter per day, while Rebel Exceda, a known shade-tolerant cultivar, had a DLIm of 8.8 moles per square meter per day. When looking at the bluegrass hybrids, though, there were some discrepancies within those cultivars, ranging from low at ~1 mole per square meter per day to high at 18 moles per square meter per day, indicating some differences in shade tolerance among bluegrass species (8).
One of the physiological shade avoidance mechanisms employed by turfgrasses is leafblade elongation. When enhanced leafblade elongation occurs, it not only causes an increase in required mowing frequency but also causes uneven energy distribution within the plant, sending more energy to the leaves while taking away from root and tiller development. The plant hormone gibberellic acid (GA) is a major response factor that is involved with leaf elongation in turfgrass being grown under shade stress. Turfgrasses grown under full sun conditions typically have lower concentrations of the biologically active gibberellin GA1 and GA4 as compared to that grown under shade and thus have lower rates of leaf elongation (12).
One gene that has been found to impact the production of biologically active GA is the GA2-oxidase (GA2ox) gene (5). Overexpression of the GA2ox gene causes the 2B-hydroxylation on GA1 and GA4 to be catalyzed, producing biologically inactive GA8 and GA34 (5). The production of these inactive GA forms limits leaf elongation under both sun and shade conditions. Overexpression of GA2ox in bahiagrass (Paspalum notatum L.) and creeping bentgrass resulted in plants that were more dwarf, had increased tiller density and decreased internode extension and darker green color, all of which contributed to improved turf quality (1). Additionally, creeping bentgrass overexpressing the GA2ox gene had decreased leaf elongation and improved coverage under 70% artificial shade (11).
Scotts ProVista (PV) KBG was developed by using a variant of the GA2ox gene originally found in spinach and inserted into KBG through gene bombardment (4). By utilizing the benefits of the GA2ox gene, PV was developed to require less mowing and have darker green color and improved density. Additionally, PV was also genetically altered to contain a variant of the 5-enolpyruvy 1-3-phosphoshikimate (EPSPS) gene to allow PV to have tolerance to the non-selective herbicide glyphosate (4).
We hypothesize that with the overexpression of GA2ox, PV will have decreased leaf elongation, darker green color and improved canopy density with improved quality under shade conditions and lower light requirements as compared to conventional KBG. To test this hypothesis, a multiyear field study was developed to quantify the DLIm and compare the shade performance of PV KBG (PV) and non-transformed KBG (CONV).
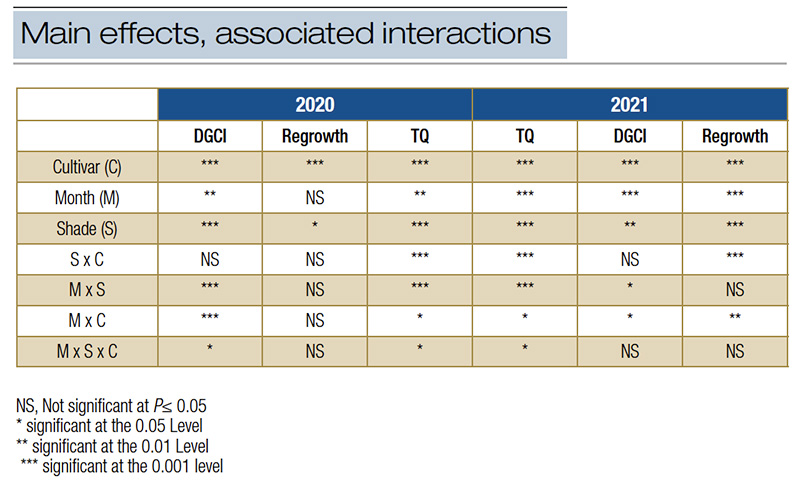
Table 1. ANOVA table for all main effects and the associated interactions of cultivar, month and shade on turf.
Materials and methods
This multiyear trial was conducted at the Scotts Miracle-Gro Research and Development Center, Marysville, Ohio, from 2019 to 2021. Research plots were established on Oct. 9, 2019, using mature sod of both GA2ox-transformed PV and non-transformed premium KBG. The cultivar composition of the conventional sod used was 30% Abbey, 30% Gaelic, 30% Jumpstart and 10% Avalanche. All sod was grown on-site before being harvested for trial use. Soil texture and classification [Pewamo silty clay loam (fine, mixed, active, mesic Typic Argiaquolls)] were similar in both locations with a soil pH of 6.5 and 3% organic matter.
All plots were fertilized using 43.72 pounds nitrogen per acre (49 kilograms nitrogen per hectare) of 32% nitrogen-0% phosphorus-4% potassium (Turf Builder, Scotts Miracle-Gro, Marysville, Ohio) per application. Applications were made at install and in mid-November in 2019 and in April, June, late August and late October during 2020 and 2021. Propiconazole 1-[2-(2,4Dichlorophenyl)-4-propyl-[1,3] dioxolan-2-ylmethyl]-1H-[1,2,4] triazole (Propiconazone 14.3 Select, Prime Source, Ankeny, Iowa) was applied every 21 days at a rate of 2.90 pounds per acre active ingredient (3.25 kilograms per hectare active ingredient) from April to October. Additionally, imidacloprid (1-[(6-chloro-3-pyridinyl) methyl]-N-nitro-2-imidazolidinimine) was applied in June and August at a rate of 1.34 pounds per acre (1.5 kilograms per hectare).
To maintain plot integrity, preemergent and post-emergent herbicides were used as needed, and irrigation was applied via overhead irrigation as needed to prevent wilt. Mowing was performed weekly at a height of 3 inches (7.6 centimeters) using a rotary mower with clippings being collected.
A strip-plot design with cultivar (PV or CONV) as the main plot (49.21 feet x 19.69 feet/15 meters x 6 meters) and the shade level as the subplot was used for this study. Five levels of shade were evaluated: full sun (0% shade), 30%, 50%, 70% and 90%. Shade structures were designed based off previous research (13) and were covered with density neutral black poly fiber shade cloths (International Greenhouse Co., Danville, Ill.) and positioned 11.81 inches (30 centimeters) above each plot. Shade was imposed from mid-June until mid-October each year and was removed once a week for data collection and plot maintenance.
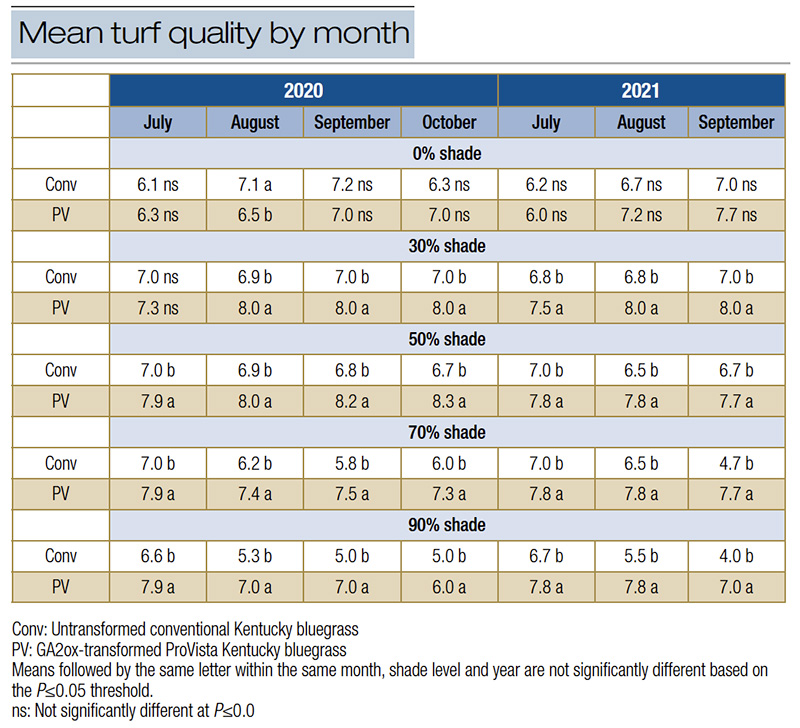
Table 2. Cultivar x Month x Shade interaction for mean turf quality across month by shade level for conventional and ProVista Kentucky bluegrass during 2020 and 2021. No data was collected during October of 2021.
Data collection
Plots were evaluated weekly from July to October each year for turf quality (TQ), dark green color index (DGCI) via light box image analysis and canopy regrowth. Visual TQ ratings were made following a modified National Turfgrass Evaluation Program 1-to-9 scale, where 7 was minimally acceptable, 1 was completely brown and 9 was perfect turf (9). DGCI was determined by taking images of each plot using a Canon Powershot G1X 14-megapixel digital camera (Canon USA) set to 1200-x-1600-pixel resolution mounted inside a 2 x 2 x 2-foot (0.6 x 0.6 x 0.6-meter) metal light box illuminated by four compact fluorescent bulbs. All images were analyzed for DGCI using turf analyzer software (http://turfanalyzer.com) (6). Finally, canopy regrowth was measured using the floating disk method and measured the difference in canopy height between weekly mowing events (10). Values for each respective trait were pooled together within each month over the duration of both years.
Photosynthesis photon flux (PPF) (micromoles per square meter per second) was also recorded under each shade treatment on five-minute intervals for the duration of the shade periods to quantify DLI. To measure the PPF, quantum light sensors (LightScout Quantum Light Sensors, Spectrum Technologies, Aurora, Ill.) were attached to data loggers (Watchdog 1000 Series Micro Stations, Spectrum Technologies, Aurora, Ill.) and mounted under each shade structure. PPF measurements from each shade treatment were summed up at the end of each year and converted to DLI values for each treatment (moles per square meter per day).
To determine the DLIm of each cultivar, turf quality from the final rating date of each year (early October and late September 2020 and 2021 respectively) was used to reflect the final quality of the stand after a full season of shade. For each cultivar, the DLIm was determined for each replication using regression of the DLI data and the final TQ ratings with a minimally acceptable TQ rating threshold of 7. The best fitting regression model selected was the quadratic linear regression model TQ= B1DLI + B2[DLI]2 + yo.
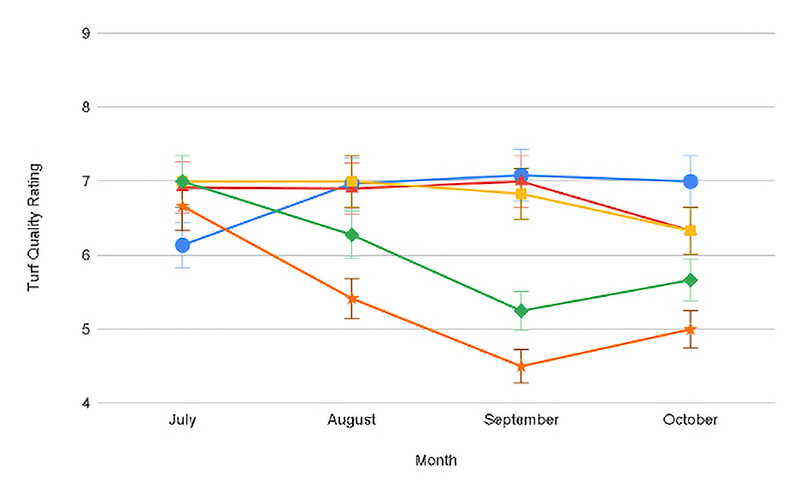
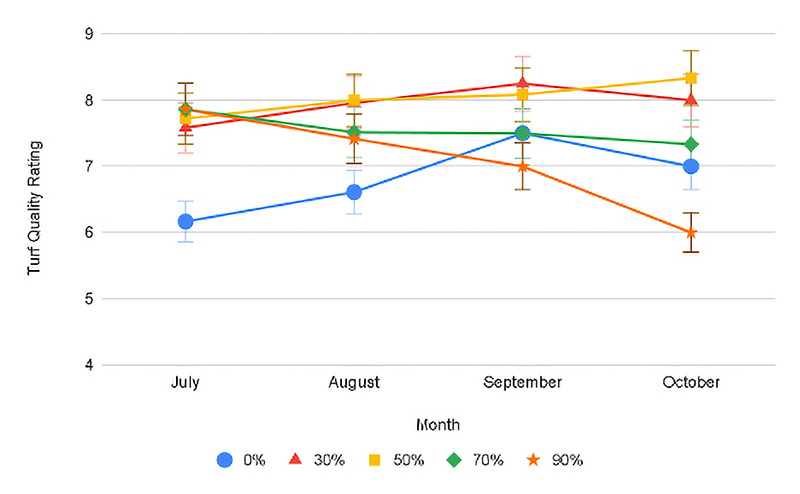
Figure 1. Progression of turf quality through the duration of shade exposure averaged across both years for conventional (top) and ProVista (bottom) KBG. Error bars represent Fisher’s LSD at P≤0.05.
Statistical analysis
All data analysis was performed using R (R core team, 2014) utilizing a repeated measures analysis of variance (ANOVA). For the DLIm analysis, a linear mixed model was used, with shade and cultivar as between-subject and dates as within-subject factors. For DGCI and canopy regrowth analysis, a linear mixed model was also used with the cultivar and shade levels as fixed effects and date as the random effect. Fisher’s Least Significant Difference (LSD) test was performed using a P≤0.05 threshold for treatment means separations.
Results
Turf quality
In both years of the trial, there was a significant month x shade x cultivar interaction for turf quality (Table 1). Both years under full sun conditions, the turf quality during July declined below the acceptable threshold of 7 to between 6.0 and 6.3. The decline was due to stand thinning and chlorosis as a result of summer heat stress under the full sun, a stress not present under shade. Except in July 2020, PV displayed significantly greater turf quality across all shade levels as compared to conventional KBG and maintained acceptable TQ (Table 2, Figure 1). Under lighter shade (30% and 50%), PV TQ remained high (7.3 and 8.3), while conventional KBG was significantly lower (6.5 and 7.0). As shade levels intensified, the quality of both cultivars decreased, but PV maintained quality ratings ≥7 in all months (7.0-7.9) except October 2020 (6.0). July was the only month that conventional KBG had acceptable quality under most shade levels. Except July, TQ ranged between 4.0 and 6.6 for conventional KBG. The quality decline under denser shade was driven primarily by decreased color, more vertical growth and thinner stands not observed in the PV. ProVista did not experience the typical shade decline common in KBG and maintained high performance under all shade levels. It appears the transformation of the GA2ox gene does confer significant shade tolerance improvement in KBG, as the performance of PV under 90% shade to our knowledge is the highest level of performance of any KBG under that shade level and is comparable to that of more shade-tolerant cool-season species.
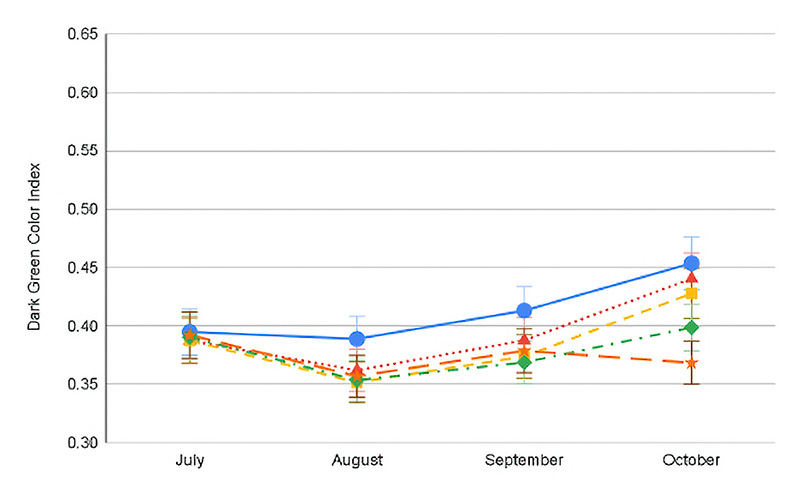
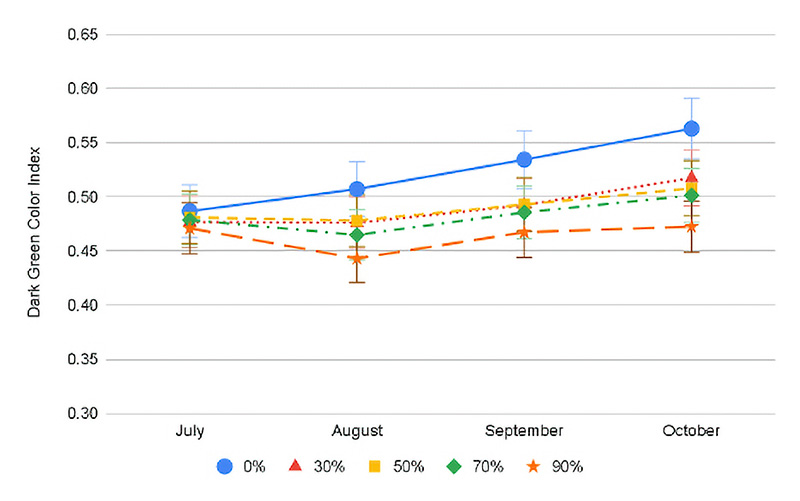
Figure 2. Progression of Dark Green Color Index value for the Month x Shade interaction averaged across both years for conventional (top) and ProVista (bottom). Error bars represent Fisher’s LSD P≤0.05
Minimum daily light integral requirements
Using the quadratic linear regression equation developed based on the results from this study, the DLIm calculated for PV was between 7.5 and 10.0 moles per square meter per day and 22.5 to 26.2 moles per square meter per day for conventional KBG in 2021 and 2020, respectively. Due to the lack of a non-transformed control, the exact benefit of the GA2ox gene cannot conclusively be determined, but the PV DLIm range is comparable to that of shade-tolerant turf-type tall fescue (8). Meeks et al. previously determined that the DLIm for Kenblue KBG was 24.1 moles per square meter per day (8), similar to that found in this study for conventional KBG. While reports are limited for the light requirements of KBG, the agreement between this study and Meeks et al. does suggest higher light requirements for KBG. Based on these results, it does appear that transformation of the GA2ox gene does significantly decrease light requirements, thus improving shade tolerance.
Dark green color index and leaf elongation
Reduced dark green color was one of the factors causing decreased turf quality under shade. Except for July in both years, full sun generally had the greatest DGCI for both cultivars (0.41-0.52), while across all the shade levels both cultivars ranged only between 0.36 and 0.42. Potential chlorophyll degradation and mild moisture or heat stress may be occurring during July under full sun exposure, which could be the reason behind the DGCI decline. In 2020, across dates and shade levels, PV had a greater DGCI (0.45-0.55) as compared to 0.35-0.45 for conventional KBG (Figure 2). Regardless of the year or cultivar, as shade levels increased, the DGCI decreased (Figure 2). Compared to conventional cultivars in this study and other studies, PV always had a greater DGCI value regardless of conditions, validating that the GA2ox gene does also improve color (1, 7).
As the shade levels got more dense, the stand density decreased, causing a decrease in quality. One of the factors that cause turf stands to become thin under shade stress is the rapid elongation of the leaf blades. As the leaves elongate more, energy is pulled away from other portions of the plant, causing stressed, unhealthy plants and decreasing stand density. Elongation rates were quantified for each level of shade by recording the turf height between each mowing event. Although regrowth rates were different between years, generally across all the shade levels, the conventional KBG (CONV) regrew on average 5-7 centimeters per week, while PV regrew only approximately 1 centimeter per week (Figure 3). The effect of shade was minimal on PV regrowth as compared to CONV, as PV showed only a 0.6% increase in regrowth rate between full sun and 90% shade, while the CONV showed a 121% increase. Regardless of shade level, CONV regrowth was significantly greater under any shade level as compared to full sun, while no difference was observed in PV. Plants that allocate less energy in shoot growth have been found to have better root and crown health and thus better plant health (14). The GA2ox gene transformation is likely the reason for the decreased leaf elongation under normal conditions and under shade stress. A similar response has been observed in creeping bentgrass transformed with the GA2ox gene, where leaf elongation was decreased under shade environments (15).
The objective of this study was to determine the DLIm and the effect on shade tolerance for KBG transformed with the GA2ox gene. Results from this study show that PV with the GA2ox gene has a lower DLIm requirement (7.5-10 moles per square meter per day) as compared to CONV (22.5-26 moles per square meter per day). ProVista also has significantly improved shade tolerance with greater turf quality, higher DGCI value and decreased leaf elongation. While a non-transformed control was not used in this study, results indicate the overexpression of GA2ox may improve shade performance of KBG.
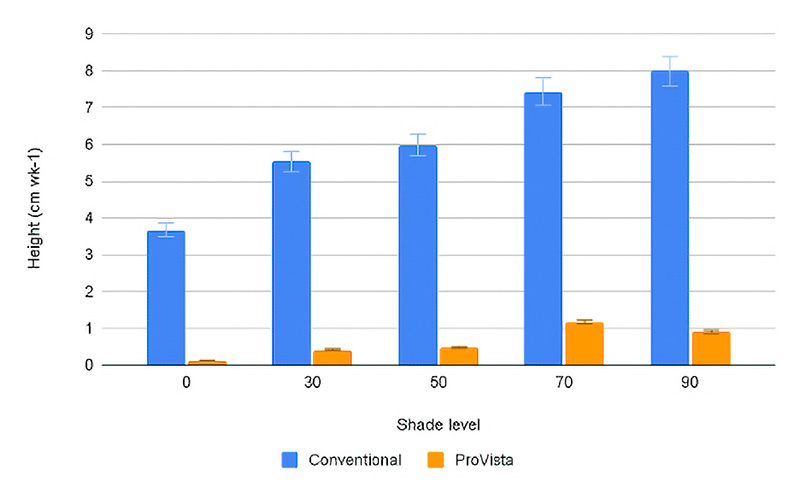
Figure 3. Weekly regrowth in centimeters of conventional and ProVista KBG averaged across both years across the five different shade levels. Error bars represents Fisher’s LSD P≤0.05.
The research says
- Compared to conventional Kentucky bluegrass, Scotts ProVista Kentucky bluegrass has significantly improved shade tolerance with greater turf quality, higher DGCI value and decreased leaf elongation.
- As shade levels intensified, the quality of both cultivars decreased, but ProVista Kentucky bluegrass maintained quality ratings ≥7 in all months (7.0-7.9) except October 2020 (6.0).
- While reports are limited for the light requirements of Kentucky bluegrass, the agreement between this study and Meeks et al. does suggest higher light requirements for Kentucky bluegrass. Based on these results, it does appear that transformation of the GA2ox gene does significantly decrease light requirements, thus improving shade tolerance.
Acknowledgements
This research was made possible with the support from the Scotts-Miracle Gro Co.
Literature cited
- Agharkar, M., P. Lomba, F. Altpeter, H. Zhang, K. Kenworthy and T. Lange. 2007. Stable expression of AtGA2ox1 in a low-input turfgrass (Paspalum notatum Flugge) reduces bioactive gibberellin levels and improves turf quality under field conditions. Plant Biotechnology Journal 5(6):791-801 (https://doi.org/10.1111/j.1467-7652.2007.00284.x).
- Beard, J.B. 1973. Turfgrass: Science and culture. Prentice-Hall, Englewood Cliffs, N.J.
- Beard, J.B. Shade stress and adaptation mechanisms of turfgrasses. International Turfgrass Society Research Journal 8:1186-1195.
- Harriman, R., L. Lee, D. Stalker and R. Torisky. 2019. Plants comprising events PP009-401, PP009-415, PP009-469, compositions, sequences, and methods for detection thereof. (Patent Number 10,501,753). U.S. Patent and Trademark Office.
- Hedden, P., and W.M. Proebsting. 1999. Genetic analysis of gibberellin biosynthesis. Plant Physiology 119(2):365-370 (https://doi.org/10.1104/pp.119.2.365).
- Karcher, D.E., and M.D. Richardson. 2003. Quantifying turfgrass color using digital image analysis. Crop Science 43(3):943-951 (https://doi.org/10.2135/cropsci2003.9430).
- Law, Q.D., C.A. Bigelow and A.J. Patton. 2016. Selecting turfgrasses and mowing practices that reduce mowing requirements. Crop Science 56(6):3318-3327 (https://doi.org/10.2135/cropsci2015.09.0595).
- Meeks, M., A. Chandra and B.G. Wherley. 2015. Growth responses of hybrid bluegrass and tall fescue as influenced by light intensity and trinexapac-ethyl. HortScience 50(8):1241-1247 (https://doi.org/10.21273/hortsci.50.8.1241).
- Morris, K. 2004. National Turfgrass Evaluation Program — information. www.ntep.org. (http://www.ntep.org/information3.htm).
- Patton, A.J., and R.C. Braun. 2020. Measurement of turf height and growth using a laser distance device. Crop Science 61(5): 3110-3123 (https://doi.org/10.1002/csc2.20295).
- Studzinska, A.K., D.S. Gardner, J.D. Metzger, D. Shetlar, R. Harriman and T.K. Danneberger. 2012. The effect of PcGA2ox overexpression on creeping bentgrass (Agrostis stolonifera L): Performance under various light environments. HortScience 47(2):280-284 (https://doi.org/10.21273/HORTSCI.47.2.280).
- Tan, Z.G., and Y.L. Qian. 2003. Light intensity affects gibberellic acid content in Kentucky bluegrass. HortScience 38(1):113-116 (https://doi.org/10.21273/HORTSCI.38.1.113).
- Trappe, J.M., D.E. Karcher, M.D. Richardson and A.J. Patton. 2011. Shade and traffic tolerance varies for bermudagrass and zoysiagrass cultivars. Crop Science 51(2):870-877 (https://doi.org/10.2135/cropsci2010.05.0248).
- Wherley, B.G., D.S. Gardner and J.D. Metzger. 2005. Tall fescue photomorphogenesis as influenced by changes in the spectral composition and light intensity. Crop Science 45(2):562-568 (https://doi.org/10.2135/cropsci2005.0562).
- Yan, J., K. Danneberger, D. Gardner, L. Lee, J. Metzger and G.-L. Wang. 2007. Effects of gibberellin 2-oxidase, phytochrome B1, and BAS1 gene transformation on creeping bentgrass photomorphogenesis under various light conditions (https://etd.ohiolink.edu/acprod/odb_etd/ws/send_file/send?accession=osu1168027982&disposition=inline).
Kirtus Houting (kirtus.houting@scotts.com) is a research specialist for biotechnology and genetics and Matthew Koch is director of biotechnology, genetics and seed, both at Scotts Miracle-Gro Co., Marysville, Ohio; Cassie Reed is a sales representative for turf and ornamentals at Gowan Company, Yuma, Ariz.; Christian Baldwin is global development manager at The Aquatrols Company, Paulsboro, N.J.; David Gardner is a professor of turfgrass science in the Department of Horticulture and Crop Science, Ohio State University, Columbus; Baoxin Chang is a post-doctoral researcher and Ben Wherley is a professor of turfgrass science and entomology, both in Soil and Crop Sciences, Texas A&M University, College Station; and Robert Harriman is owner at Bob Harriman Consultant LLC, Delaware, Ohio.